

Nuevas publicaciones-otras entidades
Mediante un lenguaje sencillo, aquí mostramos las reseñas de algunas publicaciones recientes sobre la biodiversidad de la Macaronesia. Se hará una excepción cuando el contenido de un trabajo, desarrollado en este caso fuera de dicho ámbito geográfico, sea de interés por su relación con algún aspecto importante de la ecología o la conservación en islas. Agradecemos a las personas que firman estos artículos su amabilidad a la hora de redactar sinopsis y aportar material fotográfico para ilustrarlas.
EL PINZÓN AZUL DE GRAN CANARIA

Carrascal, L.M., A. Delgado, V. Suárez & Á.C. Moreno (2022). Population size, abundance, habitat relationships and the result of a translocation programme in the Gran Canaria Blue Chaffinch Fringilla polatzeki. Bird Conservation International 32: 460-475.
Ecología y resultados de las traslocaciones del pinzón azul de Gran Canaria. En general, existe un alto grado de taxones endémicos en ambientes insulares a lo largo de todo el mundo. Con frecuencia, especies de aves insulares cuentan con áreas de distribución y poblaciones reducidas, lo que las hace muy vulnerables a impactos humanos como la destrucción del hábitat o la introducción de depredadores foráneos. Por añadidura, las poblaciones de estas especies tienden a recuperarse con menor robustez que sus parientes continentales. A consecuencia de todo ello, la mayor parte de las extinciones de aves en los últimos cuatro siglos se ha producido en islas oceánicas.
El pinzón azul de Gran Canaria (Fringilla polatzeki) es un paseriforme endémico de la isla de Gran Canaria cuya población principal se distribuye por la Reserva Natural Integral de Inagua (38 km²; figura 1). Recientemente se ha logrado establecer una segunda población en el pinar de La Cumbre (20 km²; foto 1) como consecuencia de un programa de reintroducción que ha empleado aves nacidas en cautividad y ejemplares silvestres traslocados desde la población de Inagua. El objetivo de este trabajo se centró en estimar el tamaño poblacional, los cambios temporales recientes y la distribución espacial de este pinzón en la Reserva Natural Integral de Inagua. También estimamos su tamaño poblacional en La Cumbre con el objeto de generar información que sirviese de base para evaluar la efectividad del programa de reintroducción en este pinar tras 10 años desde su comienzo.

Figura 1. La Reserva Natural Integral de Inagua se sitúa al oeste de la isla de Gran Canaria, abarcando los pinares de Inagua, Ojeda y Pajonales, todos ellos maduros y desarrollándose en condiciones ambientales semiáridas. En la foto insertada, un macho de pinzón azul de Gran Canaria (Fringilla polatzeki) en las cercanías de una fuente natural presente en esta reserva integral.
Los muestreos en la Reserva Natural Integral de Inagua y en los pinares de La Cumbre se realizaron mediante una red de transectos lineales divididos en unidades de 500 m que se repitieron en tres ocasiones entre los meses de mayo y junio de cada año (2017-2019). La abundancia de pinzones en Inagua se relacionó con la disponibilidad del hábitat modelada con las características ambientales del entorno de nidos exitosos en ese pinar publicados con anterioridad. Además, se generó un mapa de la variación espacial en Inagua considerando la abundancia de la especie estimada en los censos.
La mayor parte de la población de Inagua estaba concentrada en los sectores altos de la reserva. Las variables que más afectaron a la distribución del pinzón azul en este pinar fueron la precipitación durante el verano, la posición geográfica latitudinal y la radiación solar en junio. De esta manera, la abundancia fue muy baja en zonas de la reserva con precipitaciones en verano por debajo de los 13 mm, aumentando rápidamente hasta los 17 mm. La abundancia de la especie aumentó con el incremento de la latitud (de sur a norte) y disminuyó con una radiación solar por encima de los 7,4 kWh/m².
Las variables relacionadas con la vegetación tuvieron poca importancia en su distribución espacial. No existió una asociación uniforme entre la disponibilidad de hábitat y la abundancia de pinzones en Inagua, de manera que en este pinar existen zonas consideradas adecuadas, pero sin su presencia o en densidades muy bajas. Esto sugiere que el pinar de Inagua no está ocupado en su totalidad, por lo que existe hábitat potencial en este enclave para albergar más ejemplares.

Foto 1. Los pinares de La Cumbre son fruto de las repoblaciones forestales realizadas a mediados del siglo pasado. Estos jóvenes pinares, situados en el centro de la isla y a mayor altitud que los pinares de Inagua, Ojeda y Pajonales, albergan una pequeña población de pinzón azul de Gran Canaria establecida como resultado de un programa de reintroducción.
La densidad promedio de la especie en Inagua en 2019 fue de 10,2 aves/km² (95% CI: 7,2-13,7), el tamaño poblacional estimado fue de 362 aves (95% CI: 257-489), y su distribución era heterogénea a lo largo de la reserva. En La Cumbre se estimó una población de 68 pinzones azules (95% CI: 35-141), registrándose una densidad promedio de 3,3 aves/km² (95% CI: 1,7-6,8). Estas densidades no solo son las más bajas registradas para aves forestales en el archipiélago canario, sino también para las que habitan en todo el Paleártico Occidental.
En conjunto, se estimó un tamaño poblacional de 430 pinzones en 2019. El 16% de la población se encuentra en el pinar de La Cumbre, donde los esfuerzos de conservación llevados a cabo por el Cabildo de Gran Canaria, con el apoyo de la Unión Europea (LIFE14 NAT/ES/000077), han logrado con éxito aumentar su área de distribución y el número de efectivos (foto 2). El resultado del programa de reintroducción de pinzones azules en La Cumbre refuerza la importancia de este tipo de acciones de conservación en la prevención de extinción de especies amenazadas, especialmente de aquellas cuyas poblaciones son muy pequeñas y restringidas a una sola localidad aislada. Alejandro Delgado (CRFS, Tafira) y Luis María Carrascal (MNCN-CSIC), mayo de 2022.

Foto 2. El establecimiento de la población de La Cumbre ha sido posible gracias a las traslocaciones de ejemplares. En la imagen, una hembra de pinzón azul de Gran Canaria nacida en cautividad ceba a su pollo en el entorno de La Cumbre tras haberse reproducido con éxito.
EMPALAMIENTO DE REPTILES EN GRAN CANARIA

Caballero, J.M., A. Balmori-de la Puente, T. Calderón, I. de la Calle & A. Balmori (2021). Southern grey shrike as a predator of reptiles on the island of Gran Canaria and a comparison between island and mainland predation rates. Herpetological Bulletin 157: 16-20.
Reptiles en la dieta del alcaudón real: comparación de las tasas de depredación de Gran Canaria con las del continente. El alcaudón real (Lanius meridionalis) es un paseriforme que se extiende por diferentes partes del suroeste de Europa y áreas discontinuas del norte de África, Oriente Medio y las zonas septentrionales del subcontinente indio. En Canarias está representado por una subespecie endémica (L. m. koenigi), que se reproduce en las islas e islotes centro-orientales de Tenerife, Gran Canaria, Fuerteventura, Lobos, Lanzarote, La Graciosa y Alegranza (foto 1).

Foto 1. En este alcaudón real (Lanius meridionalis) se puede observar la ausencia de un pico y garras robustos, aspecto que lo diferencia de las verdaderas rapaces diurnas. Esta carencia ha sido compensada con el desarrollo de un refinado comportamiento a fin de manejar presas grandes y consumirlas de manera fraccionada: el empalamiento.
Pese a que la dieta de esta especie ha sido ampliamente estudiada en diversas regiones del mundo, incluyendo algunas islas del archipiélago canario, hasta la publicación de este trabajo no había información sobre dicho aspecto ecológico en Gran Canaria. Asimismo, sabemos que las islas Canarias presentan unas condiciones ambientales propias, en las que los reptiles han evolucionado hasta dar lugar a una gran diversidad de endemismos, y donde las aves constituyen uno de sus principales depredadores. Además, debido a la estabilidad de las condiciones climáticas y la disponibilidad de reptiles durante todo el año en la islas, los alcaudones no necesitan migrar a otras zonas durante el invierno como puede ocurrir en el continente. Pero, ¿qué importancia tiene la depredación de los alcaudones sobre estos reptiles exclusivos?, y, ¿es similar el porcentaje de depredación de reptiles en islas que en ambientes continentales? El estudio, por tanto, fue llevado a cabo con el fin de 1) conocer la dieta del alcaudón real en Gran Canaria mediante sus presas empaladas, y 2) comparar, haciendo una revisión bibliográfica, la importancia de los reptiles como fuente de alimento para este paseriforme en hábitats insulares y del continente.
Con el propósito de resolver el primer objetivo del trabajo, durante los años 2015 y 2016, muestreamos un entorno costero en el municipio de Agüimes, sureste de Gran Canaria, bordeando la Zona de Especial Conservación (ZEC) ES7010052 Punta de la Sal. La finalidad del muestreo era localizar empalamientos llevados a cabo por el alcaudón real y así describir su dieta en esa isla. Durante 21 jornadas de campo, se identificaron y determinaron a nivel específico 64 empalamientos, pudiéndose comprobar que un porcentaje elevado de las presas (89%) fueron reptiles endémicos (figura 1).

Figura 1. Empalamientos de las cuatro especies de reptiles encontradas en este estudio: A) lagarto gigante de Gran Canaria (Gallotia stehlini) en sustrato vegetal de espino de mar (Lycium intricatum), B) perenquén de Boettger (Tarentola boettgeri) en sustrato artificial, C) lisa grancanaria (Chalcides sexlineatus) en sustrato vegetal de espino de mar, y D) lagarto atlántico (Gallotia atlantica) en sustrato artificial.
En cuanto al segundo objetivo, que conllevó la comparación del índice de depredación en islas con el del continente, fueron consultados 22 estudios (incluyendo el presente) donde se proporciona información sobre este aspecto de la dieta del alcaudón real, empleando distintos métodos de muestreo (empalamientos, examen de egagrópilas y presas aportadas al nido) y formas de análisis, como por ejemplo el porcentaje y biomasa aportada por los reptiles con respecto al total de presas. Los resultados de la revisión indican que el alcaudón real es un animal oportunista, que basa su dieta en una gran diversidad de familias de reptiles en función de su disponibilidad en las distintas localidades estudiadas.
En los estudios hechos a partir de egagrópilas, los reptiles, debido a su mayor tamaño, conforman un más alto porcentaje en biomasa que en número de individuos en sí, incluyendo los invertebrados. Los resultados también indican que al considerar los empalamientos el porcentaje de reptiles que aparece es mayor que en otros métodos de muestreo, como el análisis de egagrópilas. Por último, su espectro trófico en las islas incluye una mayor abundancia de reptiles que en los continentes, lo que resalta la importancia ecológica de este tipo de presas para esta y otras especies de la avifauna insular. José M. Caballero y col., noviembre de 2021.
CONTROL DE COTORRAS INVASORAS

Saavedra, S. & F.M. Medina (2020). Control of invasive ring-necked parakeet (Psittacula krameri) in an island Biosphere Reserve (La Palma, Canary Islands): combining methods and social engagement. Biological Invasions 22: 3653-3667.
Control de la cotorra de Kramer en la isla de La Palma. Durante los últimos tiempos, el medio canario ha venido soportando el afincamiento de numerosas especies exóticas invasoras, entre las que se incluyen aves tan notorias y problemáticas como el miná común (Acridotheres tristis), el bulbul de vientre rojo (Pycnonotus cafer), la cotorra argentina (Myiopsitta monachus) y la cotorra de Kramer (Psittacula krameri) (foto 1).

En cuanto a paseriformes concierne, el miná común ya se considera erradicado de toda Canarias, una vez confirmada su desaparición de Tenerife en 2000, de Gran canaria en 2006 y de Fuerteventura en 2010. Respecto al género Pycnonotus, correspondiente a los bulbules, está representado por dos especies que han incidido en mayor o menor medida sobre ciertos componentes de la fauna nativa de sendas islas. En Tenerife, por ejemplo, una población reproductora de bulbul de cachete rojo (P. jocosus) fue detectada y eliminada con éxito en el año 2007, mientras que el bulbul de vientre rojo pudo ser constatado viviendo en libertad en Fuerteventura durante 2010. Desde entonces, esta especie se ha dispersado de manera considerable, ocupando hoy día casi toda la vertiente oriental de la isla, una franja notablemente antropizada que va desde Corralejo hasta Morro Jable.
Si hablamos de las psitácidas, la repartición de la cotorra argentina en el archipiélago canario comprende Gran Canaria y los sectores meridionales de Tenerife (año 2015) y Fuerteventura (2018). Por el momento, está ausente de La Palma (2015-2020) y no ha llegado a establecerse en La Gomera, ni tampoco en El Hierro (2015), mientras que en la isla de Lanzarote no hay datos actualizados, excepto algunos avistamientos esporádicos (2015).
Foto 1. La cotorra de Kramer (Psittacula krameri), junto con otros loros y córvidos, es un magnífico ejemplo de ave particularmente inteligente, de ahí la dificultad en el trampeo de sus poblaciones introducidas.
Por su parte, la cotorra de Kramer está presente en Tenerife y Gran Canaria, islas donde ha ido ampliando su rango de distribución (2015). En Fuerteventura, sin embargo, se encuentra en franca regresión, sobre todo a decir por el carácter residual de su población reproductora actual, quizá formada por unos cinco ejemplares avistados en Morro Jable (2018). Para La Gomera y El Hierro hay algún que otro registro de la especie en el pasado, pero, que sepamos, no se ha producido asentamiento (2015). Tampoco existen datos actualizados para Lanzarote, salvo ciertas observaciones ocasionales (2015). Un caso aparte son las dos poblaciones reproductoras que había en la isla de La Palma (una en la zona de Breña Alta y otra, más reducida, en Los Llanos), puesto que fueron sometidas a acciones de manejo cuyo resultado fue la eliminación de todos los individuos existentes en el medio natural. Estos trabajos anularon los efectos negativos de esta cotorra invasora en la biota insular, principalmente en lo que atañe a la repercusión de su actividad en las especies nativas, tanto vegetales, de las que se alimentaba, como aviares, con las que competía. Además, su eliminación fue crucial para el sector primario, más aún al tener en cuenta, según estudios previos, el nefasto efecto que ejercían sobre más de 22 cultivares de la isla.
Todas las labores de seguimiento y control de la cotorra de Kramer en La Palma fueron desarrolladas priorizando el consenso con la ciudadanía y, por consiguiente, minimizando el conflicto social. A día de hoy, el Cabildo Insular de La Palma tiene operativo, dentro del marco del proyecto 2015/2018, un Equipo de Rápida Respuesta compuesto por voluntariado especializado. Gracias a su actuación, una vez finalizada la campaña inicial, ya han sido capturados varios ejemplares recientemente liberados, neutralizando por tanto su expansión por el medio natural de la isla. Los avisos de la población local, que ha seguido los protocolos consensuadamente establecidos, ponen de manifiesto un alto grado de concienciación en lo que a este programa de eliminación de especies invasoras se refiere. Susana Saavedra (INBIMA) y Félix M. Medina (Cabildo de La Palma e IPNA-CSIC), noviembre de 2020.
LOTUS LEÑOSO EN LA GOMERA

Portero Álvarez, A.M., J. Martín-Carbajal González, J.A. Reyes-Betancort & R. Mesa Coello (2019). Lotus gomerythus (Fabaceae-Loteae) spec. nova. Botánica Macaronésica 30: 89-98.
Lotus gomerythus, una nueva fabácea (Loteae) para Canarias. Aquel día de febrero de 2016, nuestra única intención era realizar una ruta “botánico-paisajística”, como a nosotros nos gusta llamarla, por el sureste de la isla de La Gomera. Al igual que en otras ocasiones, durante el itinerario también se contemplaba la escalada para acceder a algunos enclaves nunca antes visitados, en ese caso un pequeño roque sálico muy característico que nos quedaba de camino. Pese a advertir cuando trepábamos que era inaccesible para los herbívoros, nunca imaginamos poder encontrar tal cantidad y variedad de plantas nativas en su parte alta. La gran curiosidad, suscitada por estar ante una comunidad aparentemente prístina, hizo que deparásemos en cada una de las especies para poder identificarlas (foto 1). Así fue como descubrimos un ejemplar de Lotus que, desde un primer momento, nos pareció que se trataba de un nuevo taxón para la biodiversidad canaria.

Foto 1. Trabajos de campo en la parte alta del roque relacionados con el hallazgo, estudio y descripción de Lotus gomerythus, en esta ocasión muestreando la comunidad vegetal acompañante.
Tras varias visitas al roque, así como de estudios morfológicos y moleculares de una única planta, hoy podemos hablar de Lotus gomerythus, un nuevo pico de la sección Rhyncholotus exclusivo de La Gomera (foto 2). Se trata de un subarbusto de porte rastrero, tallos leñosos en la base, con toda la parte vegetativa de la planta vellosa, hojas sésiles, seríceo plateadas, linear-filiformes, y flores de color naranja-azafrán, con tonos ferruginosos en el estandarte. Las alas, visiblemente más cortas que la quilla y no arropando a esta, hace que las flores se diferencien claramente de las de los picos de fuego (L. pyranthus) y cernícalo (L. eremiticus) palmeros, mientras que su ápice redondeado las distingue de las de los picos de paloma (L. bethelotii) y de El Sauzal (L. maculatus) tinerfeños (figura 1).

Foto 2. Flor de Lotus gomerythus, un nuevo pico para la ciencia descubierto en La Gomera en 2016.
En lo alto del roque podemos encontrar numerosas especies vegetales que acompañan al solitario nuevo pico, entre ellas aparecen árboles achaparrados como la sabina (Juniperus turbinata subsp. canariensis) y el acebuche (Olea cerasiformis), además de otras de porte mucho más reducido como rudas (Ruta sp.), lenguas de pájaro (Globularia salicina), magarzas (Argyranthemun frutescens), tederas (Bituminaria bituminosa), veroles (Kleinia neriifolia), anís de risco (Bupleurum salicifolium), tabaibas (Euphorbia berthelotii), pataconejo (Polycarpaea divaricata), tomillos gomeros (Micromeria gomerensis), cerrajas (Sonchus sp.), cardoncillos (Ceropegia dichotoma subsp. krainzii), lavandas (Lavandula canariensis subsp. gomerensis), férulas (Ferula linkii), gamonas (Asphodelus ramosus), verodes (Aeonium decorum y A. canariense subsp. latifolium), tajinastes (Echium aculeatum), balillos (Atalanthus canariensis), tasaigos (Rubia fruticosa) y scillas (Scilla latifolia), entre otras. Observar tremendo elenco en este reducto escarpado nos hace pensar en cómo pudo haber sido la corología y densidad de todas estas especies en un pasado lejano, antes de que fuesen drásticamente restringidas por la fuerte presión antrópica, mediada sobre todo por los ganados en rincones como este.

Figura 1. Muestra de flores de (A) pico de paloma (Lotus berthelotii), (B) L. gomerythus spec. nova y de (C) pico de El Sauzal (L. maculatus) en las que se aprecian claramente las diferencias de color, tamaño y forma de sus alas y cálices.
El hallazgo de una única planta de L. gomerythus evidencia ya el grave peligro de extinción en el que se encuentra. Urge ahora su pronta inclusión en los catálogos de especies amenazadas y adoptar, de manera paralela, medidas preventivas de choque para evitar tener que lamentar su pérdida, entre ellas, la clasificación del lugar donde se encuentra como zona de exclusión y la búsqueda de más ejemplares en otros riscos gomeros. Ana Portero y col., diciembre de 2019.
ARAÑAS DEL GÉNERO DYSDERA EN CANARIAS

Macías-Hernández, N., S. de la Cruz López, M. Roca-Cusachs, P. Oromí & M.A. Arnedo (2016). A geographical distribution database of the genus Dysdera in the Canary Islands (Araneae, Dysderidae). ZooKeys 625: 11-23.
Base de datos sobre la distribución geográfica del género Dysdera en las islas Canarias. Las arañas del género Dysdera han protagonizado un gran proceso de radiación adaptativa en el archipiélago canario. Hasta el momento se conocen en las islas unas 47 especies endémicas, las cuales están incluidas en un total de 250 cuya distribución abarca la Macaronesia, Marruecos atlántico y la cuenca mediterránea. Son arañas nocturnas de mediano tamaño (figura 1), que se encuentran principalmente en hábitats de nuestras islas caracterizados por la humedad, incluyendo además el medio subterráneo o cavernícola.
En este trabajo se presenta una base de datos con la distribución actualizada de todas las especies del género presentes en Canarias (un total de 794 registros documentados desde 1971 hasta el 2015), así como los mapas de distribución georreferenciados de cada una de ellas.
Figura 1. Algunas de las especies de Dysdera presentes en las islas Canarias: D. calderensis (a), D. longa (b), D. verneaui (c) y D. unguimmanis (d).

El número de especies de cada isla depende de factores biogeográficos como el área, la altitud, la diversidad de hábitats, la edad y la lejanía. La combinación idónea favorece a las islas centrales (de edad intermedia), que disponen de una mayor riqueza específica, mientras que las islas más jóvenes y alejadas (La Palma y El Hierro), así como las más antiguas y erosionadas (Lanzarote y Fuerteventura), albergan el menor número (figura 2). La gran riqueza de especies en Canarias hace que muchas de ellas se encuentren simultáneamente en un mismo hábitat, de modo que las que coexisten tienden a ser de tamaños distintos y con sus partes bucales de diferente morfología. Esto hace pensar en la probable segregación trófica entre especies de Dysdera para evitar la competencia por los recursos alimenticios.
Figura 2. Representación gráfica del número de especies de Dysdera presentes en las islas Canarias. Negro: número de especies compartidas con otras islas; gris: endemismos locales. Las líneas conectoras representan el número de especies compartidas entre islas, y el grosor de las mismas es proporcional a la cantidad de especies compartidas. La disposición de los círculos refleja la localización geográfica de las islas: La Palma (P), El Hierro (H), La Gomera (G), Tenerife (T), Gran Canaria (C), Fuerteventura (F) y Lanzarote (L).
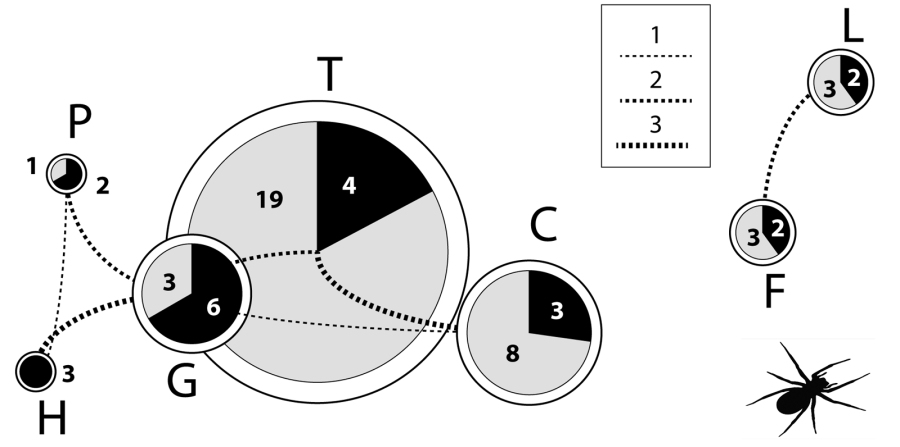
La base de datos georreferenciada y los mapas presentados en esta publicación serán de gran utilidad para futuros estudios biogeográficos, taxonómicos, ecológicos y de conservación. Nuria Macías-Hernández (Dep. Biol. Anim. & Edafol. Geol.-ULL) y col., enero de 2017.
DISPERSIÓN SECUNDARIA DE SEMILLAS POR RAPACES

López-Darias, M. & M. Nogales (2016). Raptors as legitimate secondary dispersers of weed seeds. Ibis 158: 428-432.
Rapaces como legítimas dispersoras secundarias de semillas. Si bien las rapaces no han sido usualmente consideradas como buenas dispersoras de semillas, en este artículo se demuestra que su papel ecológico en los mecanismos de dispersión de plantas herbáceas podría ser relevante. En la isla de Fuerteventura, el busardo ratonero o aguililla (Buteo buteo) y el cernícalo vulgar (Falco tinnunculus) dispersan secundariamente (esto es, dispersan semillas consumidas por sus presas) un sinfín de semillas de diversas especies. Hasta 11.000 semillas, de 39 y 62 especies distintas de plantas herbáceas respectivamente, fueron encontradas en aproximadamente 300 egagrópilas analizadas de cada rapaz. Estas semillas fueron consumidas directamente por especies invasoras en la isla, como conejos (Oryctolagus cuniculus) y ardillas morunas (Atlantoxerus getulus), que forman parte importante de la dieta de estas dos aves rapaces (figura 1). De las especies de plantas encontradas en las egagrópilas de aguililla y cernícalo, cuatro y siete respectivamente aparecieron en más de un 10% de las egagrópilas. De éstas, algunas mostraron viabilidad en su germinación, incluso después de haber pasado por el tracto digestivo de los herbívoros que primero las consumieron y de las rapaces que depredaron posteriormente sobre estos. Este estudio inspira un cambio de perspectiva en el papel de las rapaces como dispersoras de semillas, y propone que éstas podrían estar involucradas en mecanismos complejos de dispersión secundaria de semillas a una escala más global, incluyendo sus movimientos potenciales a larga distancia. Marta López-Darias y Manuel Nogales (IPNA-CSIC), julio de 2016.

Figura 1. A: Ejemplar adulto de una de las rapaces objeto de estudio, el busardo ratonero (Buteo buteo), conocido comúnmente en el archipiélago canario como “aguililla” y cuyo papel en la dispersión secundaria de semillas de plantas herbáceas se muestra relevante. B: Individuo de ardilla moruna (Atlantoxerus getulus), un roedor introducido en Fuerteventura en 1965 y que en la actualidad está distribuido abundantemente por toda la isla; esta especie, junto con el conejo, también invasor, constituye una de las principales presas de las aves rapaces estudiadas y son los consumidores o dispersores primarios de las semillas de plantas herbáceas. C: Detalle de la flor de cosco (Mesembryanthemum nodiflorum), una de las plantas herbáceas que es secundariamente dispersada por estas rapaces.
ORIJAMAS Y LAGARTOS GIGANTES CANARIOS

Pérez-Méndez, N., P. Jordano & A. Valido (2015). Downsized mutualisms: Consequences of seed dispersers’ body-size reduction for early plant recruitment. Perspectives in Plant Ecology, Evolution and Systematics 17: 151-159.
Efectos de la reducción del tamaño de los dispersores de semillas en el reclutamiento temprano de plantas. La reducción del tamaño de los vertebrados frugívoros mediada por la acción humana puede conllevar una pérdida de la calidad de los servicios de dispersión de semillas que estos ofrecen. Como consecuencia, la regeneración y la dinámica de las poblaciones de plantas que dependen de ellos pueden verse afectadas negativamente. En este trabajo evaluamos las consecuencias de la extinción y reducción del tamaño corporal de los lagartos gigantes de Canarias (gén. Gallotia, Lacertidae) en la regeneración natural de la orijama o leña buena (Neochamaelea pulverulenta, Rutaceae), un arbusto endémico de las islas que depende exclusivamente de dichos reptiles para dispersar sus semillas (figura 1).

Figura 1. Individuo adulto de orijama (Neochamaelea pulverulenta) con numerosos frutos maduros (A). Estos frutos son consumidos exclusivamente por lagartos endémicos de las islas Canarias (gén. Gallotia) de talla mediana o grande. Su eficacia como dispersores de semillas de dicha planta va en función del tamaño que alcanzan: (B) el lagarto de Lehrs (G. caesaris, La Gomera) es un dispersor muy poco eficiente debido a su pequeña talla (longitud hocico-cloaca máxima [en adelante, máx. LHC] = 111 mm); (C) el lagarto tizón (G. galloti, Tenerife) es un dispersor sub-eficiente por su talla mediana (máx. LHC = 145 mm); y (D) el lagarto gigante de Gran Canaria (G. stehlini) es un dispersor eficiente debido a su gran talla (máx. LHC = 280 mm).
Nuestros resultados no muestran diferencias en los patrones de estructura de edad (componente cuantitativa), es decir, las poblaciones con lagartos grandes mostraron una proporción similar de individuos de distintas clases de edad (plántulas, brinzales, juveniles y adultos) que las poblaciones con lagartos medianos y pequeños. Sin embargo, sí se detectó una reducción significativa de la tasa de reclutamiento efectiva, es decir, el número de individuos creciendo fuera de la copa de plantas adultas, y del vigor de las plántulas (componentes cualitativas) en las poblaciones que albergan lagartos de tamaño medio y pequeño. Este trabajo resalta la importancia de conservar a los vertebrados frugívoros de mayor tamaño y todo el rango de procesos funcionales (componente cuantitativa y cualitativa) envueltos en las relaciones mutualistas para mantener la regeneración y dinámica poblacional. Néstor Pérez-Méndez (EBD-CSIC) y col., mayo de 2015.
DESCRIPCIÓN DE UNA NUEVA FABÁCEA CANARIA

Gil González, J., J.B. Morales Mateos, M.L. Gil González & R. Mesa Coello (2013). Vicia voggenreiteriana (Fabaceae) a new species from the island of La Gomera (Canary Islands). Vieraea 41: 189-201.
Descrita una nueva planta exclusiva de La Gomera. Al hallazgo en 2010 de la fabácea Vicia vulcanorum en Lanzarote, un nuevo y sorprendente endemismo de esta isla, se suma en esta ocasión el de otra especie del mismo género en La Gomera. Su descripción científica, en el último número de la revista Vieraea, está a cargo del etnobotánico Jaime Gil, del arqueobotánico Jacob B. Morales y de los botánicos Ricardo Mesa y Manuel L. Gil.
Foto 1. Inflorescencia de Vicia voggenreiteriana. Nótese especialmente la acusada longitud del pedicelo floral y del tubo del cáliz.

Vicia voggenreiteriana (foto 1) es el nombre que se ha dado a este endemismo gomero como tributo al Dr. Volker Voggenreiter (1941-2002), botánico alemán que dedicó una buena parte de su vida profesional al estudio de la flora y vegetación de nuestro archipiélago. Está localizado únicamente en el sur de la isla, concretamente en el barranco de Guarimiar (Alajeró), donde uno de los autores (RM) recolectó las primeras semillas en 2010, justo cuando realizaba un seguimiento de las poblaciones insulares de trébol de risco blanco (Dorycnium eriophthalmum) en los acantilados de La Galga. Fue precisamente la particular apariencia de dichas semillas, muy diferente a las del resto de especies endémicas del género Vicia en Canarias, lo que motivó su inmediato estudio (figura 1). Con el fin de comprobar la estabilidad de sus caracteres, fue sembrada una muestra en Lanzarote bajo unas condiciones edafoclimáticas muy diferentes a las de la localidad de origen. Las plantas resultantes de dicha siembra mantenían las mismas características y mostraban caracteres florales y vegetativos también diferentes. De manera paralela, y dada la práctica ausencia de especímenes de herbario de este género procedentes de La Gomera en los principales herbarios canarios y peninsulares, se emprendió una campaña recolectora específica a fin de aportar el material necesario para concluir que nos encontrábamos ante una nueva planta y afrontar tanto su descripción original como su comparación con las especies más afines.
La descripción de esta nueva especie botánica, así como las de otras que publican diferentes autores en el mismo número de Vieraea (Tolpis santosii, Ferula arnoldiana, Sideritis soluta ssp. hildae y Sideritis santosii), pone de manifiesto la importancia que tiene el estudio de la extraordinaria biodiversidad florística de nuestras islas, además de los usos culturales asociados a las diferentes plantas, un legado que estamos obligados a preservar para las futuras generaciones. Jaime Gil González (Dep. Cienc. Hist., ULPGC) y col., enero de 2014.

Figura 1. Vista ventral de las semillas de Vicia de Canarias. De izquierda a derecha, V. voggenreiteriana n. sp. (Guarimiar, La Gomera), V. scandens (Aguamansa, Tenerife), V. vulcanorum (Malpaís de La Corona, Lanzarote), V. nataliae (Vallehermono, La Gomera), V. cirrhosa (Las Eras, Tenerife) y V. sp. (Chira, Gran Canaria). La escala muestra 1 mm.
ALCATRACES Y BASURA MARINA

Rodríguez, B., J. Bécares, A. Rodríguez & J.M. Arcos (2013). Incidence of entanglements with marine debris by northern gannets (Morus bassanus) in the non-breeding grounds. Marine Pollution Bulletin 75: 259-263.
Incidencia de los enganches con basura marina en los alcatraces atlánticos. En la actualidad, una enorme cantidad de plástico se distribuye, desde la superficie hasta el fondo, a lo largo y ancho de todos los océanos del planeta. Este material, de origen antrópico, procede de fuentes muy diversas (barcos, ríos, sistemas de alcantarillado costeros, etc.) y está compuesto principalmente por restos de las artes de pesca, de material de embalaje o desechos de la industria plástica. A consecuencia de la ingestión accidental y de los enganches que se producen, toda esta basura provoca una afección muy negativa a la vida marina en general. Es bien conocido, por ejemplo, que numerosas aves marinas en todos los mares del mundo ingieren plásticos con los consiguientes problemas de salud que ello conlleva. No obstante, el efecto y la magnitud de sus enganches con pedazos de artes de pesca u otra basura es mucho más difícil de evaluar debido, seguramente, a la baja detectabilidad de los casos, así como a sesgos relacionados con el hecho de que normalmente solo son estudiados los animales ya muertos por esta causa. En el marco del proyecto INDEMARES, se trató de paliar la susodicha falta de información aprovechando los censos estandarizados de aves marinas que se llevaron a cabo. Por el mayor tamaño y fácil detección de la especie, fue evaluada la incidencia de enganches con plásticos en el alcatraz atlántico (Morus bassanus), un ave que nidifica en el Atlántico Norte y que fuera de la época de cría está distribuida por las aguas de la península ibérica (foto 1), del noroeste de África y del Mediterráneo occidental.
Foto 1. Ejemplares jóvenes de alcatraz atlántico concentrados en las costas cantábricas durante el periodo no reproductor.

Foto 2. Un alcatraz atlántico inmaduro, fotografiado en aguas de Mauritania, acarreando una cuerda enredada en su pico.

Aunque los enganches también ocurren en el mar, parece que algunos alcatraces pueden enredarse con plásticos en las colonias de cría, sobre todo cuando es conocido que regularmente estos residuos son utilizados en la construcción de sus nidos. No existía, sin embargo, información previa de lo que ocurre en el mar, fuera de las áreas de cría. Por ello, durante seis campañas oceanográficas, a bordo de buques del Instituto Español de Oceanografía (IEO) y del Instituto Social de La Marina (ISM), cubriendo la totalidad de las aguas españolas y de la plataforma del noroeste de África, fueron anotadas todas las observaciones hechas al respecto: se registraron los datos y, en la medida de lo posible, se hicieron fotografías de cada una de las aves que tenían plásticos enredados. Gracias a esto fue posible cuantificar la incidencia de los enganches en las distintas zonas de invernada, así como describir las principales características de los objetos.
Se comprobó que la incidencia de los enganches varió de forma importante entre las distintas zonas, alcanzando hasta el 20% de las aves en las aguas de Mauritania. La mayoría de los ejemplares afectados, en este caso con objetos enganchados a la mandíbula inferior, fueron inmaduros (foto 2). En este sentido, la técnica de pesca de la especie está basada en picados a gran velocidad, por lo que es posible que los enganches se produzcan al confundir los plásticos con presas, o al intentar capturar peces asociados a la basura que flota (los peces pelágicos suelen concentrarse debajo de objetos flotantes para protegerse). Buena parte de estos objetos provenía claramente de actividades como la pesca o la navegación (cuerdas y nylon), y el color predominante de los mismos fue el rojo. Al no conocer la proporción de colores en la basura marina en general, surge la duda sobre si el color rojo es seleccionado activamente o si realmente es el más abundante en el medio. Serían necesarios más estudios para dar a conocer cuál es el alcance real de estos enganches en las poblaciones de alcatraces y otras especies marinas. Su finalidad sería, además, la búsqueda de soluciones sostenibles –por ejemplo, el cambio paulatino del color de las artes de pesca– frente a un problema que se intuye grave... Beneharo Rodríguez (SEO/BirdLife-Canarias) y col., septiembre de 2013
NUEVOS CARACOLES TERRESTRES ENDÉMICOS DE LA GOMERA

Santana, J., M. Artiles, Y. Yanes, F. Deniz, M.R. Alonso & M. Ibáñez (2013). Three undescribed species of Napaeus (Gastropoda: Pulmonata: Enidae) from La Gomera (Canary Islands). The richest centre of species radiation for the genus. Journal of Conchology 41: 271–286.
Yanes, Y., J. Martín, J. Santana, G.A. Holyoak, D.T. Holyoak, M. Artiles, F. Deniz, M.R. Alonso & M. Ibáñez (2011). Four new Napaeus species (Gastropoda: Pulmonata: Enidae) from La Gomera (Canary Islands). Journal of Conchology 40: 393−407.
Descripción de nuevos caracoles terrestres del género Napaeus endémicos de La Gomera. Con una distribución que abarca todas las islas, Napaeus es el género de caracoles terrestres con mayor número de especies conocidas en Canarias. De momento, se tiene constancia de unas 68 vivientes, más una fósil de Tenerife y otra posiblemente extinta de Gran Canaria. Curiosamente, es en la isla de La Gomera, una de las más pequeñas de este archipiélago, donde más especies han sido descritas hasta ahora, en concreto 26, lo que representa más de 1/3 del total. Siete de ellas se han dado a conocer en dos trabajos publicados en fechas recientes: en el primero hacemos alusión a cuatro cuya distribución incluye la mitad norte de la isla, y en el segundo a tres colectadas en la mitad meridional (figura 1). La descripción de estas nuevas especies gomeras ha estado fundamentada en la morfología de sus conchas y la anatomía del genital. Cabe destacar que la isla alberga tanto a las especies más pequeñas como a las más grandes del género y que, además, en ellas se advierte una mayor variedad de modelos diferentes del aparato reproductor.

Figura 1. Algunas de las nuevas especies de caracoles terrestres del género Napaeus halladas en la isla de La Gomera (islas Canarias): N. doloresae (A-B), N. estherae (C) y N. magnus (D) (lámina publicada en Santana y col., 2013).
Un aspecto interesante de Napaeus es que muchas de sus especies pueden disfrazarse de manera activa, adhiriendo suelo y/o líquenes a su concha: bien revolcándose en la tierra húmeda, formando una cubierta más o menos uniforme, o recogiendo con la rádula (boca) partículas de suelo o de líquenes y pegándolas con saliva (foto 1). Esta habilidad les ayuda a pasar desapercibidos ante sus depredadores naturales. Yurena Yanes (Dep. Geol.-UC) y col., septiembre de 2013

Foto 1. Individuo activo de Napaeus gomerensis en cuya concha se aprecia una costra de protuberancias de líquenes, adheridas de manera deliberada, que le sirve para pasar inadvertido sobre el sustrato rocoso.
NUEVA ESPECIE DE ERIZO DE MAR

Rodríguez, A., J.C. Hernández, S. Clemente & S.E. Coppard (2013). A new species of Diadema (Echinodermata: Echinoidea: Diadematidae) from the eastern Atlantic Ocean and a neotype designation of Diadema antillarum (Philippi, 1845). Zootaxa 3636: 144–170.
Diadema africanum, un conocido erizo de mar y una nueva especie para la ciencia. Esta investigación pone de manifiesto que Diadema africanum, el erizo diadema o de Lima, muy “popular” en Canarias por ser el principal responsable de originar los blanquizales (foto 1), muestra diferencias genéticas y morfológicas suficientes como para separarlo de su paisano, el distribuido por el Atlántico oriental, D. antillarum. Al comparar los ejemplares procedentes de Madeira, Canarias y Cabo Verde con el aquí también designado neotipo de D. antillarum (espécimen que sirve de tipo nomenclatural a falta de otro material que fundamente el nombre del taxón), así como con ejemplares del Caribe, advertimos claramente dichos contrastes.
Foto 1. Tras el intenso ramoneo de Diadema africanum, las rocas quedan desprovistas de macroalgas y paulatinamente van adquiriendo el color blanquecino característico de las algas calcáreas.

Además de la distancia genética, las diferencias morfológicas que dan validez a esta nueva especie son la presencia de un solo tipo de pedicelario tridentado (apéndice con función limpiadora, defensiva, etc.), unas púas más estrechas y cortas, un patrón de iridióforos (foto 2; células que refractan o reflejan la luz) bien marcado y arcos genitales pronunciados en su sistema apical. Su distribución, por tanto, comprende la plataforma continental de Senegal y Ghana, así como las islas oceánicas de Cabo Verde, São Tome, Canarias, Salvajes y Madeira. El resultado de este estudio nos hace ver que el erizo al que habíamos clasificado durante mucho tiempo como introducido es en realidad autóctono, una especie bien diferenciada que ha habitado estos fondos marinos desde épocas remotas. Adriana Rodríguez (BIOECOMAC-ULL) y col., julio de 2013

Foto 2. Detalle del patrón de iridióforos del sistema apical de Diadema africanum.
AUTILLOS EXTINTOS DE MACARONESIA

Rando, J.C., J.A. Alcover, S.L. Olson & H. Pieper (2013). A new species of extinct scops owl (Aves: Strigiformes: Strigidae: Otus) from São Miguel Island (Azores Archipelago, North Atlantic Ocean). Zootaxa 3647: 343–357.
Rando, J.C., H. Pieper, J.A. Alcover & S.L. Olson (2012). A new species of extinct fossil scops owl (Aves: Strigiformes: Strigidae: Otus) from the Archipelago of Madeira (North Atlantic Ocean). Zootaxa 3182: 29–42.
Figura 1. Reconstrucción del autillo de Madeira realizada por Pau Oliver, y representación esquemática de esta misma especie y del autillo europeo en la que se observa una gran diferencia entre la longitud de las patas de estas aves.

Los avances en el conocimiento de las biotas originales de los archipiélagos oceánicos, gracias al estudio de sus registros paleontológicos, indican que las islas poseían una riqueza y diversidad mucho más alta que la actual. Las faunas insulares actuales son pobres subconjuntos de las originales. En la mayoría de los casos, este empobrecimiento comenzó con la llegada de población humana a los archipiélagos. Las islas macaronésicas son un claro ejemplo de estos procesos y las investigaciones realizadas en este campo muestran la existencia de fuertes eventos de extinción entre los vertebrados.
Nuestros últimos trabajos describen a dos de estas aves, el autillo de Madeira (Otus mauli) y el autillo de São Miguel (O. frutuosoi), endémicas de los archipiélagos de Madeira y Azores, respectivamente. Los restos de Madeira ponen de manifiesto que se trataba de una especie con unas alas de talla similar a las del autillo europeo (O. scops), aunque con unas patas más largas y estilizadas (figura 1) y un peso algo superior. Estas características apuntan a una vida ligada al suelo, probablemente en el sotobosque de la laurisilva de esa isla.
La especie de São Miguel tenía unas alas más cortas (14%; figura 2), pero unas patas más largas (11%) que el autillo europeo. Su peso estimado fue también más de un 30% superior. Estas características y las proporciones de sus huesos indican unos hábitos terrestres y menos voladores que el autillo europeo (figura 3), pero similar a los de otros búhos también extintos de Hawaii (género Grallistrix) y Nueva Zelanda (Sceloglaux albifacies).

Figura 2. Ala derecha del autillo europeo y tamaño estimado del ala del autillo de São Miguel, ambas con los huesos en posición anatómica correspondientes a cada especie. La diferencia entre las dos superficies alares es de un 33%. Escala = 4 cm.
La extinción de ambas especies parece estar asociada a la llegada humana a sus respectivos archipiélagos y a las alteraciones posteriores, como la introducción de roedores o gatos, depredadores a los que pudieron ser especialmente vulnerables. La datación 14C de algunos de los huesos de la especie de São Miguel ofrece una edad de unos 2.000 años (49 cal BC–125 cal AD), por lo que su extinción tuvo que ser posterior a esta fecha. Juan Carlos Rando y col., junio de 2013

Figura 3. Huesos de las patas (a: fémur; b: tibiotarso; c: tarsometatarso) del autillo de São Miguel (a-c), del autillo de Madeira (a`-c`), y del autillo europeo (a``-c``), donde se aprecia la mayor longitud de los de las especies de Macaronesia, especialmente en los tarsometatarsos. Escala = 2 cm.
DISPERSIÓN SECUNDARIA DE SEMILLAS

Padilla, D.P., A. González-Castro & M. Nogales (2012). Significance and extent of secondary seed dispersal by predatory birds on oceanic islands: the case of the Canary archipelago. Journal of Ecology 100: 416-427.

Foto 1. El alcaudón real es un paseriforme de hábitos depredadores. En las islas Canarias, este pequeño carnívoro basa su dieta en lagartos, los cuales representan más del 70% de la biomasa total que ingiere. Esto supone que actúe como un dispersor secundario de semillas muy efectivo, puesto que la mayor parte de las que pasan por su tracto digestivo son capaces de permanecer con una alta viabilidad y capacidad de germinación. En ocasiones, los lagartos capturados son empalados por las propias aves en espinos, retamas y otras especies de plantas, siendo este un comportamiento exclusivo de la familia de los alcaudones.
Importancia de la dispersión secundaria de semillas por aves depredadoras en islas oceánicas: el caso de las islas Canarias. La dispersión secundaria de semillas ocurre cuando dos o más dispersores intervienen y por lo general amplían la distancia a la que éstas son usualmente dispersadas. Este proceso ecológico ha sido constatado en las islas Canarias, donde aves depredadoras, como el alcaudón real (Lanius meridionalis, foto 1) o el cernícalo vulgar (Falco tinnunculus), se alimentan de lagartos (Gallotia spp.) frugívoros e indirectamente dispersan las semillas que con anterioridad habían consumido estos mismos reptiles (foto 2).

Foto 2. La saurocoria o dispersión de semillas por reptiles es un proceso eminentemente insular. Esto es debido a que los reptiles de ambientes insulares sufren una menor depredación y competencia, ya que muchos de los depredadores y competidores que tendrían en el continente son incapaces de colonizar las islas. Así, los lagartos amplían su espectro alimenticio accediendo a recursos que en el continente serían consumidos por otras especies. Un claro ejemplo de esto ocurre en las islas Canarias, donde los lagartos endémicos del género Gallotia incluyen hasta más de un 50% de frutos en su dieta.
En el presente estudio, donde fueron evaluadas las islas y los hábitats (foto 3) potenciales para que se produjese el fenómeno de la dispersión secundaria de semillas, aparecieron 78 especies de plantas diferentes en el análisis de 2.098 egagrópilas de alcaudones y 5.304 de cernícalos. Se observó cómo los cernícalos dispersaron un mayor número de plantas que los alcaudones (76 y 26, respectivamente). Del total de las plantas identificadas, el 73% fueron clasificadas como nativas, mientras que el resto (27%) concernía a especies introducidas, algunas de ellas con carácter claramente invasor. La mayor parte de las plantas detectadas en la dispersión secundaria presentaron frutos carnosos y procedían principalmente de hábitats abiertos cercanos a las zonas costeras. Los resultados del presente trabajo muestran cómo la dispersión secundaria de semillas en las islas, además de jugar un papel fundamental en los sistemas de dispersión a larga distancia, puede considerarse un proceso común y generalizado. David P. Padilla (IPNA-CSIC y UEA) y col., mayo de 2013

Foto 3. Algunas especies vegetales del Parque Nacional del Teide (Tenerife) están altamente amenazadas y luchan contra la extinción. Tanto el cedro canario como el rosal del guanche son ejemplos de plantas con poblaciones relegadas a pequeños reductos muy fragmentados. En el presente estudio, éstas y otras especies también incluidas en los listados de la UICN han sido detectadas en los sistemas de dispersión secundaria de semillas. Dicho proceso de dispersión a larga distancia podría permitir a estos taxones en peligro de extinción la colonización de nuevas zonas, además de facilitar la conexión entre poblaciones aisladas.
ALIMOCHES Y HUMANOS EN SOCOTRA

Gangoso, L., R. Agudo, J.D. Anadón, M. de la Riva, A.S. Suleyman, R. Porter & J.A. Donázar (2013). Reinventing mutualism between humans and wild fauna: insights from vultures as ecosystem services providers. Conservation Letters 6: 172-179.

Foto 1. Los alimoches son respetados y muy apreciados por los socotríes, quienes valoran el importante servicio ecosistémico que estos proporcionan. En la imagen vemos un grupo de alimoches alimentándose apaciblemente en un wady (palabra árabe que denomina el lecho de un arroyo que solo lleva agua superficial intermitentemente) en las afueras de Qalansiyah, donde niñas socotríes acaban de tirar la basura.
Mutualismo relíctico entre alimoches y humanos en Socotra. La milenaria relación mutualista entre humanos y buitres ha desaparecido de la práctica totalidad del planeta, en paralelo a los drásticos cambios económicos y sociales ocurridos en las sociedades modernas y al precipitado declive de las poblaciones de carroñeros ocurrido a nivel global. Afortunadamente, aún quedan algunos refugios donde todavía perdura esta fructífera convivencia, como el caso de los alimoches comunes o “guirres” (Neophron percnopterus) en la isla índica de Socotra (foto 1). Para describir esta relación mutualista, analizamos los factores que determinan la distribución espacial de los alimoches y cuantificamos la proporción de restos orgánicos de origen humano que estas aves consumen (foto 2). Pero el desarrollo ha llegado hasta este remoto paraíso y un importante conflicto entre la modernización y la conservación de la biodiversidad se cierne sobre el futuro de la isla y sus habitantes. Laura Gangoso (EBD-CSIC) y col., febrero de 2013

Foto 2. La población de alimoche socotrí, al igual que todas las poblaciones insulares de la especie, es sedentaria, y alcanza una densidad de 0,22 parejas/km², cifra formidable si la comparamos con las 0,05 parejas/km² estimadas para la segunda población más densa, la de la península ibérica, o la de otras poblaciones insulares como Canarias (0,02 parejas/km²) o Sicilia (0,0003 parejas/km²). La estrecha y pacífica relación alimoches-humanos es manifiesta en Socotra. Los alimoches se adentran diariamente en las zonas urbanas en busca de alimento. La capital de la isla, Hadiboh, acumula enormes cantidades de basura que es depositada directamente en las calles. Además, dos restaurantes depositan cada día los restos de la cocina en un barril, siendo eficazmente consumidos por los alimoches en el transcurso de pocas horas.

© 2013 Grupo de Ornitología e Historia Natural de las islas Canarias